Von der Einzelligkeit zur Mehrzelligkeit bei Algen
Chlamydomonas
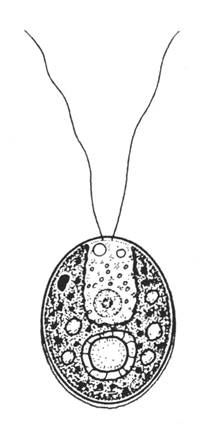
Typisch ist der becherförmige Chloroplast von Chlamydomonas, der ein grosses Pyrenoid enthält und den Zellkern umgibt. Im vorderen Bereich des Chloroplasten, dicht unter Plastidenhülle und Plasmalemma liegt das karotinoidhaltige Stigma. Der eigentliche Photorezeptor wird aber von den peripher dazu liegenden Membranen gebildet.
Die vorn gelegenen isokonten Geisseln durchdringen eine typische Wandpapille. Sie schlagen synchron nach dem „Ruderschlagprinzip“: ausgestreckt nach hinten, mit einem von der Basis zur Spitze laufenden Knick nach vorn, so wird verhindert, dass die Zellen beim Schlag nach vorn wieder zurücktreibt. Am apikalen Pol liegen ferner die pulsierenden Vakuolen, die sich alternierend füllen und entleeren.
Bei der vegetativen Fortpflanzung verläuft die Trennung der Tochterzellen vom vorderen zum hinteren Pol. Mehrere Teilungen können innerhalb der Mutterzellwand aufeinander folgen, wobei es zur Umorientierung der Achsen kommt. Nach der Begeisselung werden die Tochterzellen durch enzymatischen Abbau der Mutterzellwand freigesetzt.
Die stets einzelligen Arten aus der Gattung Chlamydomonas gehören zu den vielgenutzten Objekten pflanzlicher Grundlagenforschung. Besondere Beachtung fanden die Kompatibilitätstypen (= Paarungstypen) von ansonsten isogamen Arten (+ und – Stämme). Vermerkt sei, dass es in der Gattung Chlamydomonas neben Isogamie auch Heterogamie und Ooogamie gibt.
Zu den Molekülen, die an der Spezifität der Paarungsreaktion (Verkleben [=Agglutination] der Geisseln zweier paarungsbereiter Gameten) beteiligt sind, gehören Glykokonjugate mit alpha-glykosidisch gebundenen Mannoseresten (L. WIESE und W. WIESE, 1975).
Tetraspora
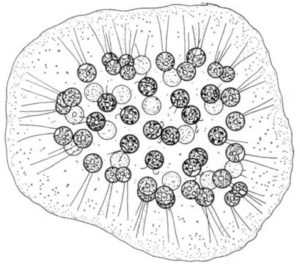 BEMERKUNG ÜBER TETRASPORALES: Manche Autoren erheben dieses Taxon zu einer eigenständigen Ordnung, andere schlagen sie den Volvocales zu. Ein Kennzeichen vieler Arten ist die Anordnung der Zellen zu Vierergruppen innerhalb einer gemeinsamen Hülle. Tetrasporales repräsentieren sicherlich keine natürliche Gruppe, denn sie sind vornehmlich ein Sammelbecken von Arten auf einer bestimmten Organisationshöhe.
BEMERKUNG ÜBER TETRASPORALES: Manche Autoren erheben dieses Taxon zu einer eigenständigen Ordnung, andere schlagen sie den Volvocales zu. Ein Kennzeichen vieler Arten ist die Anordnung der Zellen zu Vierergruppen innerhalb einer gemeinsamen Hülle. Tetrasporales repräsentieren sicherlich keine natürliche Gruppe, denn sie sind vornehmlich ein Sammelbecken von Arten auf einer bestimmten Organisationshöhe.
Gonium
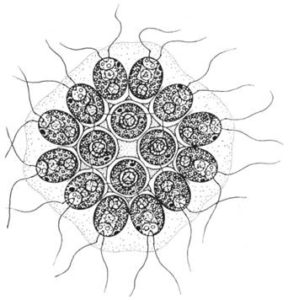 Die Gattung Gonium ist am einfachsten gebaut. Gonium sacculiferum besteht aus vier chlamydomonas-ähnlichen Zellen, die von einer gemeinsamen Gallerte umgeben sind. Die Coenobien von Gonium pectorale enthalten meist 16 Zellen, die anderer Arten 8, 16 oder 32.
Die Gattung Gonium ist am einfachsten gebaut. Gonium sacculiferum besteht aus vier chlamydomonas-ähnlichen Zellen, die von einer gemeinsamen Gallerte umgeben sind. Die Coenobien von Gonium pectorale enthalten meist 16 Zellen, die anderer Arten 8, 16 oder 32.
Pandorina
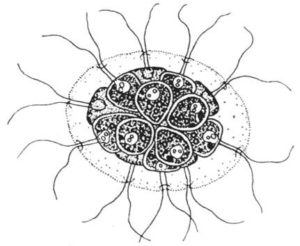 Pandorina ist das bekannteste Beispiel für Coenobienbildung. Eine Kolonie enthält auch hier wieder meist 16 (-32) Zellen, und aus jeder geht wieder eine Kolonie mit 16 (- 32) Zellen hervor usw.
Pandorina ist das bekannteste Beispiel für Coenobienbildung. Eine Kolonie enthält auch hier wieder meist 16 (-32) Zellen, und aus jeder geht wieder eine Kolonie mit 16 (- 32) Zellen hervor usw.
Der Gattungsname entstammt der griechischen Mythologie. Die Götterbotin Pandora verbreitete Unheil, und jedem Unheil folgte neues Unheil.
Eudorina
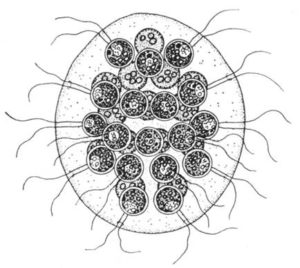
Eine ähnliche Organisation finden wir bei Eudorina. Beide Gattungen unterscheiden sich aufgrund der Anordnung und der Zahl der Zellen im Coenobium.
Pandorina-Zellen bilden einen kompakten Haufen, Eudorina-Zellen sind in Form einer Hohlkugel angeordnet.
Volvox
 Die Gitterkugel Volvox (auch Kugelalge genannt) bildet riesige Kolonien, in denen 1000 bis 50’000 Zellen hohlkugelförmig vereint sein können. Der Koloniedurchmesser beträgt bis zu zwei Millimeter. A. van LEEUVENHOEK sah sie zum ersten Male im Januar 1700.
Die Gitterkugel Volvox (auch Kugelalge genannt) bildet riesige Kolonien, in denen 1000 bis 50’000 Zellen hohlkugelförmig vereint sein können. Der Koloniedurchmesser beträgt bis zu zwei Millimeter. A. van LEEUVENHOEK sah sie zum ersten Male im Januar 1700.
Volvox-Kolonien weisen eine deutliche Differenzierung und Arbeitsteilung auf. So wird der reproduktive Bereich zunehmend auf Zellen des hinteren Pols beschränkt. Die vorderen Zellen haben oft längere Geisseln und grössere Stigmen.
Alle Zellen reagieren phototaktisch, ihre Bewegungen sind durch den Lichteinfall synchronisiert. Es gibt daher eine bevorzugte Bewegungsrichtung der Kolonie. Zwischen den Zellen findet ein Informationsaustausch statt. Der sichtbare Ausdruck hiervon ist ein symmetrisch gebautes Netzwerk aus Plasmabrücken, durch die die Zellen untereinander in Verbindung stehen. Die Kolonie reagiert stets als Ganzes.
Man könnte daher durchaus von einem echten Vielzeller sprechen. Doch die Art, wie er entstanden ist, ist kein Modell zum Verständnis der Evolution der übrigen Vielzeller. Deren Entstehung verlief anders: einfacher und effizienter.
Zur Fortpflanzung gelangen reproduktive Zellen ins Kolonieinnere, wo aus ihnen durch zahlreiche aufeinanderfolgende Teilungen nach einem charakteristischen Schema Tochterkolonien entstehen. Die Geisseln der Tochterkoloniezellen sind anfangs ins Kolonieinnere gerichtet. Durch einen „Inversions“-Prozess (Umstülpungsprozess) erlangen sie die richtige Orientierung.
Die Freisetzung der Tochterkolonien vollzieht sich nach dem Reissen der schützenden Hülle der Mutterkolonie. Dabei stirbt der Mutterorganismus ab (!). Der Modus der sexuellen Fortpflanzung ist die Oogamie.